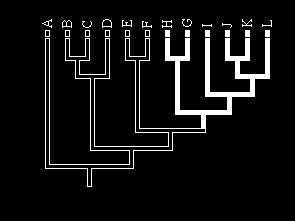
1
Phylogeny and classification
Review of approaches to classification
Classification as tool for communication
Classification as information managment
Classification as inspiration
Classification as an ideological weapon
Archaic classification
Aristotle (384-322 B.C.)
John Ray (1627-1705)
Carl Linnæus (1707-1778)
Georges Cuvier (1769-1832)
Chevalier de Lamarck (1744-1829)
'Evolutionary systematists' (app. 1910-1970)
Phylogeny (app. 1960s onwards)
Definition: hypothetical evolutionary history of organisms
Philosophical background
Techniques to construct phylogenies
Assumptions
Possibilities and limitations
Different presentations
Biology vs. palaeontology
Phylogeny as an interpretation tool
Evolutionary history
Adaptation
Biogeography
Coevolution
The new classification of classification abolished ?
2
A duck has a beak.
A crow has wings.
A herring has gills.
A duck has a beak, has wings and does not have gills.
A crow has a beak, has wings and does not have gills.
A herring does not have a beak, does not have wings but does have gills.
3
1) We use characters (properties) to classify the objects.
2) We selectively apply some and ignore other properties.
3) We allow variation even within characters.
4) Classification may or may not be hierarchic.
5) Classification is separate from taxonomy (giving names).
4
Scala natura
Aristotle (384-322 B.C.)
'Artificial' classification
Essentialism
Individuals are flawed representations of their 'essence' (ideal)
Vertebrates (with red blood)
Viviparous (giving birth to living individuals) vertebrates
Humans
Whales
Other mammals
Oviparous (egg laying) vertebrates
Birds
Amphibians and reptiles (but see snakes below)
Snakes
Fish
Invertebrates (without red blood)
Cephalopods
Crustacea
Insects, spiders.
Other mollusks, echinoderms.
Sponges, etc.
5
John Ray (1627-1705)
British botanist and vicar.
Postulated the existence of species.
Species are invariable creations of God.
There is no lack of perfection, no intermediates.
Chief ideologist of creationism.
Artificial classification (Historia Generalis Plantarum , 1686-1704) of plants based on fruits, leaves and flowers.
Artificial classification of animals, based on toes and teeth.
Carl Linnæus (1707-1778)
Son of priest, 'naturalist'.
Classified all known organisms.
Species are real, invariable entities; created constant and separate.
Artificial classification of plants by their sexual characters: (Systema Naturæ, 1735), flowering plants by the number of stamen, non-flowering by shape of female organs.
Systema Naturæ lacks embellishments.
6
Georges Cuvier (1769-1832)
Catastrophist
Multiple creations
Young Earth (thousands of years)
Fossils remains of extinct organisms
Branching classification
Rigid boundaries
Artificial classification:
Vertebrates (deer, birds, fish - anything with bones!): characterized by an internal skeleton, a brain and a spinal chord - a concentrated nervous system.
Mollusks (bivalves, gastropods, cephalopods, etc.): separate neural masses.
Articulates (lobsters, spiders, insects, some worms - animals with joints & sections): 2 ventral nerve chords, transition between circulatory system of previous and tracheae.
Radiates (star fish, sea urchins & whatever doesn't fit above): radial symmetry, no circulation, no nervous system.
Jean Baptiste Antoine de Monet,
Chevalier de Lamarck (1744-1829)
Evolutionist
Gradual evolution
Old Earth (millions of years)
Fossils remains of old specimens of living organisms
Linear classification
Fuzzy boundaries
Natural classification, fine-tuningscala natura .
7
|
'Lamarckism' sensu Darwin |
'Lamarckism' sensu Lamarck |
|
Environmental feed-back, acquired characteristics are inherited by descendants. There is a feed-back from environment to the species, through the individual.
Example: a giraffe reaches for leaves sitting higher, it gets a longer neck, and its off-spring inherits longer necks. The mechanism is an environmental interaction. |
Animals evolved from 'gelatinous' particles and plants from 'mucilaginous' particles by heat and electricity. An inner force improves species to advance up the scala natura (following Robinet). Bodies of organisms shaped by behaviour, caused by needs.
Example: giraffe gets a longer neck because it needs to reach for leaves sitting higher. Mechanism is an inner force, a need for perfection. |
Cuvier:
- species go extinct
- extinct species may be very similar to extant
- extinct species can be classified by same branching scheme
Lamarck:
- species are variable
- any classification will have to deal with intermediates
- fossils are spot-samples from a continuous history
8
9
|
Phenetics |
Phylogeny (Cladistics) |
|
R.R. Sokal & P.H.A. Sneath 1963: Principles of numerical taxonomy
'Artificial' classification Character analysis No subordination Classification reflects similarity 'Similar' = similar All characters equally good Initially very rigorous methods Distinct groups
Philosophical basis of cladistics
|
Willi Hennig (1913-1976): 1950: Grundzüge einer Theorie der phylogenetischen Systematik. 1963: Phylogenetic Systematics.
'Natural' classification Character analysis No subordination Classification reflects evolution 'Similar' = common origin All characters not equally good Initially very 'loose' methods Distinct groups
Later: biological interpretation
|
10
|
Archaic |
Scala natura |
Ray |
Linnæus |
|
|
Artificial/natural system |
- |
Artificial |
Artificial |
Artificial |
|
Organisms real or representations |
- |
Representations |
Real |
Real |
|
Categories |
Distinct |
Distinct |
Distinct |
Distinct |
|
Hybrids |
- |
- |
Do not exist |
Troublesome |
|
Linear or branching |
- |
Linear |
Linear |
Branching |
|
Characters |
Few, implicit |
Few, explicit |
Few, explicit |
Few, explicit |
|
Subordination of characters |
Yes |
Yes |
Yes |
Yes |
|
Cuvier |
Lamarck |
Thiele |
Phenetics |
Cladistics |
|
|
Artificial/natural system |
Artificial |
Natural |
Natural |
Artificial |
Natural |
|
Organisms real or representations |
Real |
Real |
Real |
Real |
Real |
|
Categories |
Distinct |
Indistinct |
Indistinct |
Distinct |
Distinct |
|
Hybrids |
Do not exist |
Evidence! |
Evidence! |
(Do not exist) |
(Do not exist) |
|
Linear or branching |
Branching |
Linear |
Branching |
Branching |
Branching |
|
Characters |
Few/some, explicit & implicit |
Few/some, explicit & implicit |
Some, mostly implicit |
Many explicit |
Many explicit |
|
Subordination of characters |
Yes |
Yes |
Yes |
No |
No (ideally) |
11
Glossary for Phylogeny
You may meet the following terms, when reading papers dealing with phylogeny. This is a 'quick & dirty' list of short explanations to help studying, not necessarily elaborate, formal definitions. I also take the liberty of explaining by rather crude examples to facilitate understanding.
Phylogeny: a) the relationship of organisms, usually presented as a branching diagram (Fig. 1) of the evolutionary pattern, and usually produced by a reproducible method (e.g., cladistics). Strictly speaking, phylogenies are merely explicit hypotheses about a relationship. Hypotheses can be tested, and may have to be falsified; what you see is the current 'best explanation of the available data', not the final truth! b) a loose term for methods and sciences contributing to the understanding of the relationship and evolution of organisms.
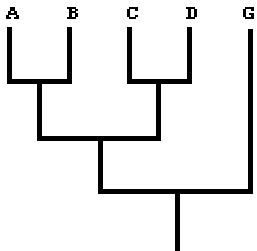
Figure 1.
Ancestral & derived: 'Ancestral' means something present in the ancestor(s), and is often used to describe a character state inferred to have evolved into a 'derived' state. In a certain context, 'fore legs' (of reptiles) are ancestral and 'wings' (of birds) derived. Note the use depends on the context. If you only study parakeets, there is no point in considering reptile legs, and the 'ancestral' character may be a feature of the general bird wing. Avoid terms like 'primitive' and 'evolved', with an implicit quality assessment. All organisms survive, and generally function very well in their environments.
Apomorphy & plesiomorphy: A plesiomorphy (plesiomorph character, plesiomorph character state) is an ancestral character state and an apomorphy (apomorph character, apomorph character state) is a derived character state. 'Wings' are an apomorphy of birds corresponding to the plesiomorph 'fore legs' of reptiles. Note the words are context specific: wings and fore legs are both apomorph 'articulated fore limbs' in the context of the phylogeny of all vertebrates (vs. plesiomorph non-articulated fins, etc.), but penguins have apomorphic 'flipper wings' relative to the plesiomorph general bird wings. Synapomorphy is a derived feature, common to all members of a clade (uniquely derived organisms) - feathered wings are a synapomorphy of birds. Autapomorphy is a derived feature found in just one member of a clade. The differences between 'apomorphy', 'synapomorphy' and 'autapomorphy' are subtle, and the terms are often used interchangeably.
Autapomorphy: See apomorphy.
Character, character states: A character is a set of homologous features - for example articulated appendages - sharing a common origin, which can be described as separate entities, character states. Character states are mutually exclusive, and every organism in the analysis can have a state assigned, and every state evolved only once. Note that character states do not necessarily describe everything conceivable, only what is relevant to your analysis! If you look at a 'bird', a 'horse' and a 'dog', the character 'articulated fore limb' may have the character states 'wing', 'hoof' and 'foot' - we totally ignore bat wings, hands, etc. They are not relevant to understand our study material, but it is essential that each state evolved only once among the organisms - they must be homologous. If we include bats in the analysis, we need to qualify the state 'wings'.
Clade: Group of organisms sharing common ancestor. The group contains that ancestor and all descendants of that ancestor. In figure 1, (A,B) is a clade, so is (C,D) and (A,B,C,D). (B,C) is not a clade; their last common ancestor had descendants, that are not in (B,C). Clades are monophyletic.
Cladistics: A particular branch of phylogenetic analysis, describing evolutionary patterns from parsimonious analysis of observed characters.
Cladogram: See tree.
Classification: Organizing organisms in a system. Formally classification, taxonomy, and phylogeny are separate issues, but many researchers do all at the same time. Note that few classifications and even less taxonomy are based on phylogeny, and no classification or taxonomy can - unless based on a phylogenetic analysis - be used for phylogenetic interpretations (interpretations of evolutionary patterns).
Derived: See ancestral.
Evolved: See ancestral.
Homology: Characters (character states) are considered homologous if they perform the same function and have the same construction, and are believed to have evolved from the same evolutionary event. Characters (character states) are considered homoplastic if they appear similar or perform similar functions, and are believed to have evolved from different evolutionary events. 'Wings' of birds are homologous - they differ, but are believed to have arisen from one event. In a larger scale 'wings', 'hooves' and 'feet' are homologous - they are different manifestations of 'articulated fore limbs', and share a common evolutionary history (vs. the absence of articulated fore limbs). But note that bird wings and bat wings are homoplastic! They perform similar functions, but evolved at separate events. Note that it is often the scope/level of resolution that determines homology.
Homoplasy: See homology.
Ingroup & outgroup: The group you are interested in analysing, for example (A,B,C,D) in figure 1 is called the 'ingroup'. In the analysis you also include an 'outgroup' (G in figure 1), that you know is more distantly related, and can be used to polarize characters.
Monophyletic: A group of organisms is monophyletic if all members have a common ancestor, the most recent common ancestor is member of the group, and all descendants of the most recent common ancestor are members of the group. In figure 1 (A,B) is monophyletic, so is (A,B,C,D), but (B,C,D) is not. (B,C,D) is paraphyletic - all members have a common ancestor, the most recent common ancestor is member of the group, but not all descendants of that ancestor are members of the group. (B,G) is polyphyletic - their most recent common ancestor is not member of the group.
Numerical taxonomy: Se phenetics.
Outgroup: See ingroup.
Paraphyletic: See monophyletic.
Parsimony (parsimonious): 'Requiring a minimal number of assumptions of events'. Explained like that, it sounds like ideal science! In phylogeny figure 1 will be considered the most parsimonious cladogram/hypothesis/interpretation if that is the arrangement of A, B, C, and D requiring the least number of evolutionary events in the characters. A and B have more in common with each other, than either has with C or D. Furthermore, for the analysis leading to figure 1 to be parsimonious, the characters should be treated as equally important. This may sound trivial, but it is not! There are other - perfectly valid - methods to develop phylogenies. Some (particularly used on DNA data) emulate the evolutionary process emulating, others employ a priori weighting of characters (some characters are treated as more important than others).
Phenetics: Originally (1960s) developed as a tool for taxonomy. Classification of organisms exclusively on 'numerical difference' (hence also known as numerical taxonomy), a calculated difference of measured or counted characters. Initially strictly non-evolutionary to avoid biases in classification, subsequently many workers softened their view to phenetic analyses reflect evolutionary history and phenetic patterns may thus depict it. Almost vanished from evolutionary biology, but techniques still used (and very valuable!) in morphometry.
Plesiomorphy: See apomorphy.
Polarity: Polarity describes the relative occurrence of character states, which are ancestral and which are derived. If a character has more than two states, polarity describes the sequence of their origin, and which evolved to which. In some cases you may define a polarity and let that be part of your analysis or you may infer it from the tree (cladogram) after the analysis. If you include an outgroup in the analysis, you define that as coinciding with the 'root' of the tree (cladogram) and thus use it to establish the polarity of characters.
Polyphyletic: See monophyletic.
Primitive: See ancestral.
Synapomorphy: See apomorphy.
Taxonomy: Strictly speaking, naming of organisms, distinct from phylogeny (establishing evolutionary pattern) and classification (organizing). Most systematists will do several or all of these in the same work, and some even claim they are three terms for the same task.
Tree: Branching diagram showing the inferred evolutionary pattern of organisms. Formally speaking, 'cladograms' and 'trees' differ by cladograms only having organisms at the tips of the branches, but trees have organisms at the nodes. Figure 1 is a cladogram as drawn, but a tree if you imagine or infer actual organisms at the branching points. Trees have a time perspective, cladograms merely depict the branching order. Biologists mostly use cladograms, palaeontologists often use trees.
12
The Phylogeny Toolbox
This bibliography gives you an entry to phylogeny, its philosophical background, techniques, and applications. Note it is biased towards cladisitics, English language and US schools of thought. There is ample literature in German and French, as well. '*' indicates personal favourites.
Philosophical background
Daniel R. Brooks & Deborah A. McLennan. 1991. Phylogeny, Ecology, and Behavior. The University of Chicago Press, Chicago & London, XII+434 pp.
Donald H. Colless. An examination of certain concepts in phenetic taxonomy. Systematic Zoology. 6-27
James S. Farris. The meaning of relationship and taxonomic procedure. Systematic Zoology. 44-51
Brian K. Hall (ed.). Homology. The hierarchical basis of comparative biology. Academic Press (New York), XVI+483 pp.
Willi Hennig. 1950. Grundzüge einer Theorie der phylogenetischen Systematik. Deutscher Zentralverlag, Berlin.
Willi Hennig. 1963. Phylogenetic Systematics. University of Illinois Press, Urbana.
David L. Hull. 1967. Certainty and circularity in evolutionary taxonomy. Evolution. 21, 174-189
Arnold G. Kluge. 1989. A concern for evidence and a phylogenetic hypothesis of relationships among epicrates (Boidae, Serpentes). Systematic Zoology. 38(1), 7-25
* John G. Lundberg & Lucinda A. McDade. 1990. Systematics. in Carl B. Schreck & Peter B. Moyle (eds.), Methods for Fish Biology, American Fisheries Society, Bethesda, XIX+684 pp. 65-108
Brent D. Mishler & Efrain De Luna. 1991. The use of ontogenetic data in phylogenetic analyses of mosses. Advances in Bryology. 4, 121-167
* Kevin C. Nixon & Quentin D. Wheeler. 1990. An amplification of the phylogenetic species concept. Cladistics. 1990(6), 221-223
* Robert J. O´Hara. 1988. Homage to Clio, or, toward an historical philosophy for evolutionary biology. Systematic Zoology. 37(2), 142-155
* Robert J. O´Hara. 1992. Telling the tree: Narrative representation and the study of evolutionary history. Biology and Philosophy. 7, 135-160
Alan de Queiroz, Michael J. Donoghue, Junhyong Kim. 1995. Separate versus combined analysis of phylogenetic evidence. Annual review of Ecology and Systematics. 26, 657-681
Kevin de Queiroz. 1985. The ontogenetic method for determining character polarity and its relevance to phylogenetic systematics. Systematic Zoology. 34(3), 280-299
* Kevin de Queiroz. 1992. Phylogenetic taxonomy. Ann. Review of Ecology and Systematics. 23, 449-480
N.R. Scott-Ram. 1989. Transformed cladistics, taxonomy and evolution. Cambridge University Press, Cambridge, XII+238 pp.
Elliot Sober. 1988. Reconstructing the past. Parsimony, evolution, and inference. MIT Press, Cambridge, XVIII+265 pp.
Phylogenetic methods & techniques
Lois A. Abbott, Frank A. Bisby & David J. Rogers. 1985. Taxonomic analysis in Biology. Computers, Models, and Databases. Columbia Universoty Press, New York. 336 pp.
* Peter Ax. 1987. The phylogenetic system. John Wiley & Sons, Chichester, XIV+340 pp.
* Kåre Bremer. 1994. Branch support and tree stability. Cladistics. 1994(10), 295-304
Daniel R. Brooks & Deborah A. McLennan. 1991. Phylogeny, Ecology, and Behavior. The University of Chicago Press, Chicago & London, XII+434 pp.
James S. Farris. The meaning of relationship and taxonomic procedure. Systematic Zoology. 44-51
Brian K. Hall (ed.). Homology. The hierarchical basis of comparative biology. Academic Press (New York), XVI+483 pp.
Willi Hennig. 1950. Grundzüge einer Theorie der phylogenetischen Systematik. Deutscher Zentralverlag, Berlin.
Willi Hennig. 1963. Phylogenetic Systematics. University of Illinois Press, Urbana.
David L. Hull. 1967. Certainty and circularity in evolutionary taxonomy. Evolution. 21, 174-189
* John G. Lundberg & Lucinda A. McDade. 1990. Systematics. in Carl B. Schreck & Peter B. Moyle (eds.), Methods for Fish Biology, American Fisheries Society, Bethesda, XIX+684 pp. 65-108
* W.P. Maddison & D.R. Maddison 1992: MacClade. Analysis of phylogeny and character evolution. Sinauer Associates, Inc., Sunderland, USA.
Alan de Queiroz, Michael J. Donoghue, Junhyong Kim. 1995. Separate versus combined analysis of phylogenetic evidence. Annual review of Ecology and Systematics. 26, 657-681
Naruya Saitou & Masatoshi Nei. 1987. The Neigbor-joining method: a new method for reconstructing trees. Molecular Biology and Evolution. 4(4), 406-425
Michael J. Sanderson. 1995. Objections to bootstrapping phylogenies: a critique. Systematic Biology. 44(3), 299-320
N.R. Scott-Ram. 1989. Transformed cladistics, taxonomy and evolution. Cambridge University Press, Cambridge, XII+238 pp.
Elliot Sober. 1988. Reconstructing the past. Parsimony, evolution, and inference. MIT Press, Cambridge, XVIII+265 pp.
D.L. Swofford & D.P. Begle 1993. PAUP, Phylogenetic Analysis Using Parsimony, Version 3.1. Users Manual, Laboratory of molecular systematics, Smithsonian Institution, Washington DC, USA
Mark Wilkinson. 1994. Three taxon statements: when is parsimony analysis also a clique analysis ?. Cladistics. 1994(10), 221-223
Quentin D. Wheeler. 1986. Character weighting and cladistic analysis. Systematic Zoology. 35, 102-109
Interpretation of phylogenies
Lois A. Abbott, Frank A. Bisby & David J. Rogers. 1985. Taxonomic analysis in Biology. Computers, Models, and Databases. Columbia Universoty Press, New York. 336 pp.
Sara S. Bretsky. 1970. Phenetic and phylogenetic classifications of the Lucinidae (Mollusca, Bivalvia). Bull. geol. Instn. Univ. Upsala N.S. 2(2), 5-23
Donald H. Colless. An examination of certain concepts in phenetic taxonomy. Systematic Zoology. 6-27
Brian K. Hall (ed.). Homology. The hierarchical basis of comparative biology. Academic Press (New York), XVI+483 pp.
Arnold G. Kluge. 1989. A concern for evidence and a phylogenetic hypothesis of relationships among epicrates (Boidae, Serpentes). Systematic Zoology. 38(1), 7-25
* John G. Lundberg & Lucinda A. McDade. 1990. Systematics. in Carl B. Schreck & Peter B. Moyle (eds.), Methods for Fish Biology, American Fisheries Society, Bethesda, XIX+684 pp. 65-108
* W.P. Maddison & D.R. Maddison 1992: MacClade. Analysis of phylogeny and character evolution. Sinauer Associates, Inc., Sunderland, USA.
Brent D. Mishler & Efrain De Luna. 1991. The use of ontogenetic data in phylogenetic analyses of mosses. Advances in Bryology. 4, 121-167
Brent D. Mishler. 1989. Review of D.L. Hull ‘Science as a Process’. Systematic Botany. 14(2), 266-268
* Robert J. O´Hara. 1988. Homage to Clio, or, toward an historical philosophy for evolutionary biology. Systematic Zoology. 37(2), 142-155
* Robert J. O´Hara. 1992. Telling the tree: Narrative representation and the study of evolutionary history. Biology and Philosophy. 7, 135-160
* Alan de Queiroz, Michael J. Donoghue, Junhyong Kim. 1995. Separate versus combined analysis of phylogenetic evidence. Annual review of Ecology and Systematics. 26, 657-681
Kevin de Queiroz. 1985. The ontogenetic method for determining character polarity and its relevance to phylogenetic systematics. Systematic Zoology. 34(3), 280-299
* Kevin de Queiroz. 1992. Phylogenetic taxonomy. Ann. Review of Ecology and Systematics. 23, 449-480
N.R. Scott-Ram. 1989. Transformed cladistics, taxonomy and evolution. Cambridge University Press, Cambridge, XII+238 pp.
Elliot Sober. 1988. Reconstructing the past. Parsimony, evolution, and inference. MIT Press, Cambridge, XVIII+265 pp.
P.F. Stevens. 1980. Evolutionary polarity of character states. Annual review of Ecology and Systematics. 11, 333-358
Phylogeny of certain groups
Sara S. Bretsky. 1970. Phenetic and phylogenetic classifications of the Lucinidae (Mollusca, Bivalvia). Bull. geol. Instn. Univ. Upsala N.S. 2(2), 5-23
C. Hedegaard. 1997. Shell structures of the Recent Vetigastropoda. Journal of Molluscan Studies. 63, 369-377
Arnold G. Kluge. 1989. A concern for evidence and a phylogenetic hypothesis of relationships among epicrates (Boidae, Serpentes). Systematic Zoology. 38(1), 7-25
* Brent D. Mishler & Efrain De Luna. 1991. The use of ontogenetic data in phylogenetic analyses of mosses. Advances in Bryology. 4, 121-167
Claus Nielsen. 1995. Animal evolution. Oxford University Press, Oxford, VI+467 pp.
J.D. Taylor, W.J.Kennedy & A. Hall. 1969. The shell structure and mineralogy of the Bivalvia, Introduction. Nuculacea-Trigonacea. Bulletin of the British Museum (Natural History), Zoology, supplement. 3
J.D. Taylor, W.J.Kennedy & A. Hall. 1973. The shell structure and mineralogy of the Bivalvia, II. Lucinacea-Clavagellacea. Bulletin of the British Museum (Natural History), Zoology. 22(9)
13
Classic examples
The rachidian is reduced in Mumbo jumboensis.
The diverse habitats of the Faraway Islands fostered a rapid speciation of Mumbidae.
A reduced osphradium in Mumbo jumboensis is an adaptation to ...
Groups with crawl-away larvae speciate more readily than those with planctonic larvae, because of limited dispersal.
The horse has a primitive stomach.
14
"Columbus sailed west from Europe and reached land in 1492"
"Columbus opened the New World in 1492"
"Columbus brought civilisation to America in 1492"
15
16
|
I |
II |
III |
IV |
|
|
A |
0 |
0 |
0 |
1 |
|
B |
0 |
0 |
1 |
1 |
|
C |
0 |
1 |
1 |
1 |
|
D |
1 |
1 |
1 |
1 |
|
I |
II |
III |
IV |
|
|
A |
0 |
0 |
0 |
1 |
|
B |
0 |
1 |
1 |
1 |
|
C |
1 |
1 |
1 |
1 |
|
D |
1 |
0 |
1 |
1 |
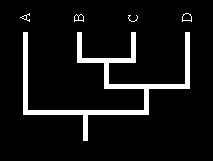
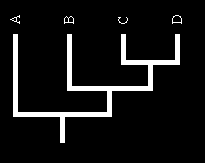
17
Characters
1. Aragonitic comarginal crossed lamellar shell structure; 2. Aragonitic radial crossed lamellar shell structure; 3. Cone complex crossed lamellar shell structure; 4. Columellar plate; 5. Operculum; 6. Propodial pedal gland; 7. Epipodium; 8. Mantle papillae; 9. Eyes; 10. Type of gills; 11. Skeletal rods; 12. Osphradium; 13. Left hypobranchial gland; 14. Right hypobranchial gland; 15. Rachidian; 16. Buccal cartilages; 17. Jaws; 18. Salivary glands; 19. Paired glands in sublingual pouch; 20. Mode of reproduction; 21. Vagina; 22. Vaginal canal; 23. Ductus enigmaticus; 24. Position of Receptaculum Seminis; 25. Spermatopore; 26. Ventricle; 27. Right auricle; 28. Position of left auricle; 29. Supraintestinal nerve.
Character diagnosis:
6. Propodial pedal gland
A distinct propodial pedal gland is present in Cocculinidae (Haszprunar, 1987), Helicinidae (Bourne, 1911; Baker, 1925, 1926), Neritopsis (Holthuis, 1995), Phenacolepadidae (Fretter, 1984; Beck, 1992), and Titiscania (Marcus & Marcus, 1967), but absent from Neritidae (Bourne, 1908), and the out-groups (Hyman, 1967). We code this character as unknown in Hydrocenidae. [0 = absent, 1 = present]
7. Epipodium.
When present in adults, the epipodium is frequently small, and may either be smooth or carry papillae/tentacles. The epipodium is absent in Patellogastropoda (Hyman, 1967), Neritilia, Neritopsis (Holthuis, 1995), and Titiscania (Bergh, 1890; Marcus & Marcus, 1967). A smooth epipodium is present in Helicinidae (Bourne, 1911), Hydrocenidae (Thiele, 1910), and the remaining Neritidae (Bourne, 1908; Holthuis, 1995). An epipodium with tentacles is present in Fissurellidae (Adams & Adams, 1858; Hyman, 1967), Cocculinidae (Haszprunar, 1987), and Phenacolepadidae (Pease, 1868; Okutani, Saito & Hashimoto, 1989; Beck, 1992). [0 = absent, 1 = present, smooth, 2 = present with tentacles]
19. Paired glands in sublingual pouch
Paired lateral glands are present in the sublingual pouches of Cocculinidae (Haszprunar, 1987), Helicinidae (Bourne, 1911; Baker, 1925), Neritidae (Thiele, 1902; Bourne, 1908), Neritopsis (Bern Holthuis, pers. comm.), and Phenacolepas (Thiele, 1902; Fretter, 1984), absent in Patellogastropoda and Fissurellidae (Hyman, 1967). From the description and position of Thiele's (1910:353) "Vorderdarmdrüsen" in Hydrocena cattaroensis, we assume these are - in spite of the unfortunate name - homologous, and tentatively assume the glands to be present in Hydrocenidae. Bergh (1890) describes the "sides of the pharyngeal anterior of the digestive tract" [our translation] of Titiscania limacina as glandular. Marcus & Marcus (1967) identify paired lateral glands in Titiscania limacina . The condition for Bathynerita, Olgasolaris and Shinkailepas is unknown. [0 = paired lateral glands absent, 1 = paired lateral glands present]
18
19
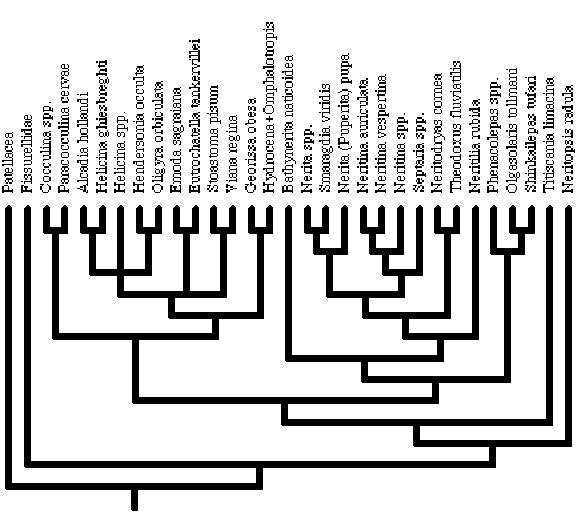
20
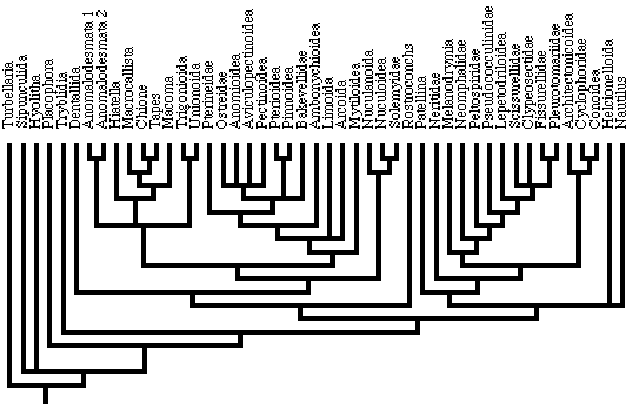
21

22

23
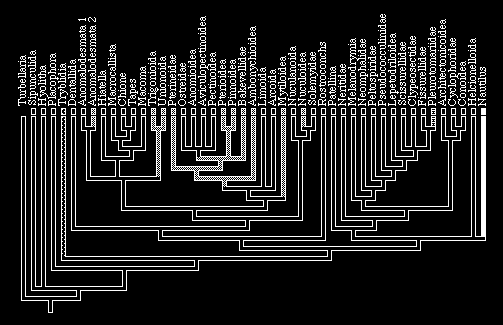
24
|
Taxon |
Geography |
Habitat |
|
A |
Mainland |
River |
|
B |
Mainland |
River |
|
C |
Mainland |
River |
|
D |
Mainland |
River |
|
E |
Island |
River |
|
F |
Island |
River |
|
G |
Island |
Lake |
|
H |
Island |
Lake |
|
I |
Island |
Lake |
|
J |
Island |
Lake |
|
K |
Island |
Lake |
|
L |
Island |
Lake |

Mainland Islands
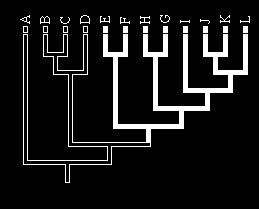
Rivers Lakes